INTRODUCTION
At first glance, an egg looks like a simple thing: a single cell, waiting to be fertilized. Yet in the fruit fly Drosophila melanogaster, making one egg is anything but simple. Behind each egg lies a temporary but remarkably complex structure called an egg chamber, built and dismantled with astonishing precision. Over the course of several days, dozens of cells organize themselves into this miniature organ, carry out a series of coordinated movements and shape changes, and then vanish, leaving behind a single, fully formed egg. How does this happen? How does one egg chamber become one egg? To answer that question, we must look inside the fruit fly ovary and follow the egg chamber from birth to disappearance, watching how individual cells collaborate to ensure that exactly one egg emerges from each chamber.
THE OVARY AS AN ASSEMBLY LINE
Fruit fly ovaries are composed of long, narrow units called ovarioles. Each ovariole functions like a conveyor belt: young egg chambers are found at one end, progressively older egg chambers appear along its length, and the most mature eggs wait at the far end to be laid.
Crucially, egg chambers do not appear fully formed. Instead, each one is assembled step by step, beginning as a cluster of cells and eventually becoming a highly organized structure capable of producing an egg. Once its job is complete, the egg chamber disappears altogether. This disposable nature makes the egg chamber an ideal system for studying development. It exists only to make one egg and nothing more.
THE GERMARIUM: WHERE EGG CHAMBERS BEGIN
Every egg chamber begins life in a specialized region of the ovariole known as the germarium. This is a crowded, dynamic environment where both germline and somatic stem cells reside.
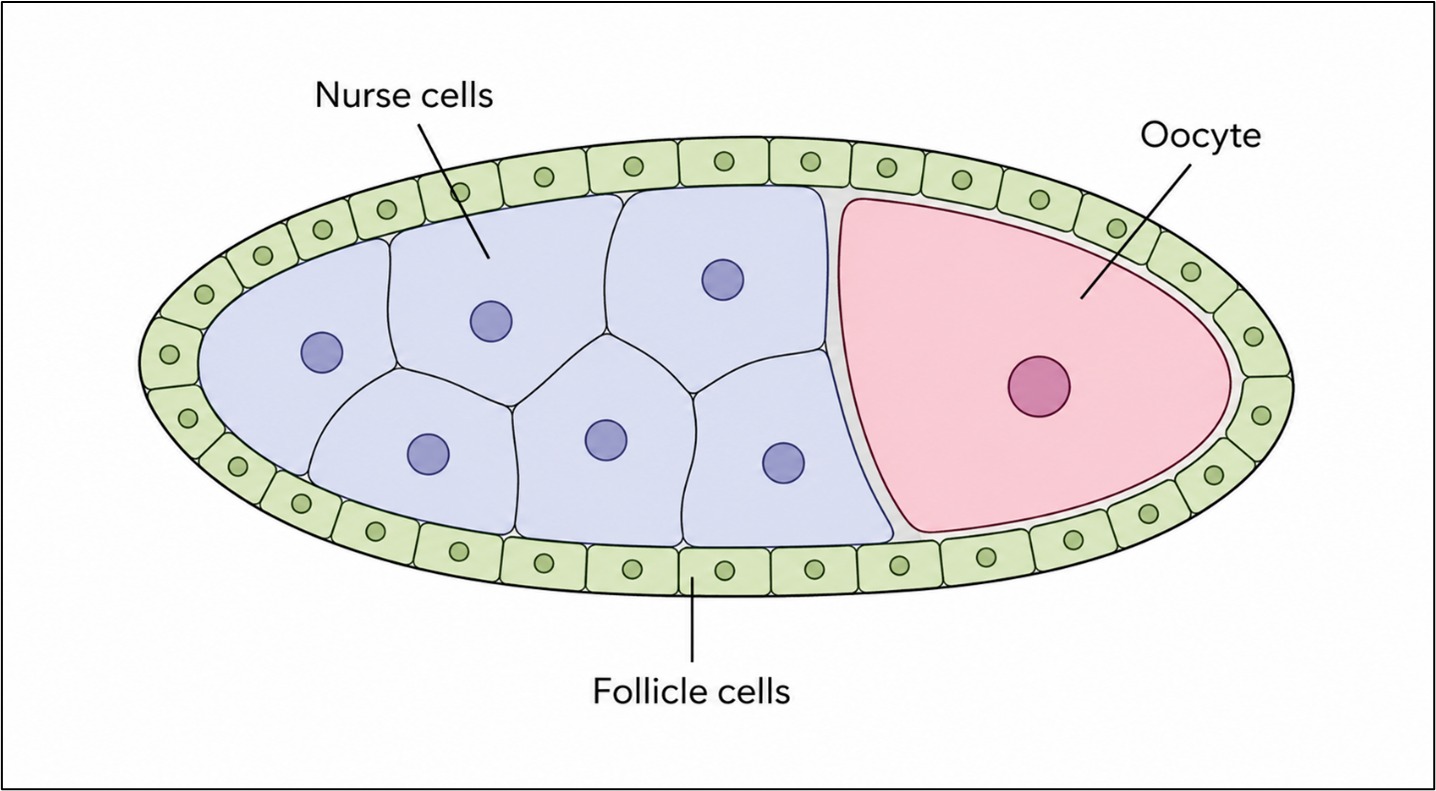
Here, germline stem cells divide to produce daughter cells that embark on a highly regulated developmental program. One daughter cell becomes a cystoblast, which undergoes four rounds of division. Importantly, these divisions are incomplete: the daughter cells never fully separate from one another. The result is a connected group of 16 germline cells, known as a 16‑cell cyst (Horne‑Badovinac & Bilder, 2005).
Although all 16 cells originate from the same parent, they quickly diverge in fate. One cell becomes the oocyte, the cell that will ultimately become the egg. Fifteen cells become nurse cells, whose sole task is to support the oocyte by producing RNA, proteins, and other materials that the oocyte alone cannot generate in sufficient quantity. At this stage, the cyst is still unprotected and indistinct from others around it. To become an egg chamber, it must be packaged and isolated.
As the 16‑cell cyst moves through the germarium, it encounters a population of somatic precursor cells. These cells are the ancestors of the follicle cells, which will later shape and protect the developing egg. These somatic cells perform a critical maneuver known as encapsulation. They extend around the cyst, migrating and wrapping until they form a continuous epithelial layer that completely surrounds the germline cells. This act physically separates the cyst from its neighbors and defines it as a distinct unit: the egg chamber.
Encapsulation is not merely a matter of coverage. It is a decisive commitment. If encapsulation fails, two cysts may end up sharing the same enclosure, leading to catastrophic developmental errors. From this point on, the fate of the egg is sealed: one cyst, one chamber, one egg.
During encapsulation, the somatic cells differentiate into three distinct types:
Follicle cells, which form the main epithelial shell
Polar cells, which anchor the chamber at its anterior and posterior ends
Stalk cells, which separate adjacent egg chambers along the ovariole
These specialized cells establish an architecture that will guide development for the rest of the egg chamber’s life (Spradling, 1993).
ARCHITECTURE OF AN EGG CHAMBER
Once encapsulation is complete, the egg chamber buds off from the germarium as a discrete structure. Its organization is simple but powerful. At its center lies the germline cyst (one oocyte and fifteen nurse cells). Surrounding it is a single layer of follicle cells, arranged as a sheet-like epithelium.
This organization creates two cooperating compartments. The germline compartment, which contains the future egg. The somatic compartment, which shapes and supports it. The follicle cell epithelium has a well-defined polarity. Each follicle cell has: An apical surface that contacts the germline and a basal surface that contacts an outer basement membrane. This polarity allows the follicle cells to behave as a coordinated tissue rather than as independent units, a property that becomes crucial during later stages when large-scale movements occur.
For a time, the egg chamber grows without dramatic rearrangements. The nurse cells begin producing materials, the oocyte slowly enlarges, and the follicle cells divide to accommodate the growing contents. Behind the scenes, however, far-reaching decisions are being made. Signals exchanged between the germline and the follicle cells pattern the epithelium along its length, dividing it into functional regions. Some follicle cells are destined to stretch, some to migrate, and others to create specialized eggshell structures. This early patterning ensures that, later on, different regions of the epithelium will behave differently, despite all follicle cells originally looking the same.
As the oocyte enters a phase of rapid growth, the egg chamber transforms. The follicle cell epithelium becomes dynamically active in a process known as epithelial morphogenesis, a collective reshaping driven by cell movements and changes in cell shape rather than by cell division.
MIGRATION OVER THE OOCYTE
Initially, follicle cells cover both the nurse cells and the oocyte. As development proceeds, most follicle cells become repositioned over the oocyte itself. This occurs through a combination of migration and dramatic changes in cell shape. Follicle cells over the oocyte become tall and columnar, packing tightly together. Follicle cells over the nurse cells become thin and squamous, stretching like living sheets to cover a large surface area. Together, these complementary transformations allow the epithelium to redistribute itself efficiently, ensuring that most follicle cells are in the right place to support the rapidly enlarging oocyte.
Even after this redistribution, the oocyte is not fully enclosed. A gap remains at its anterior edge. To close it, a subset of follicle cells performs a final, decisive movement known as centripetal migration. These cells move inward between the nurse cells and the oocyte, progressing steadily until they meet cells migrating from the opposite side. As they advance, the epithelial sheet seals shut, fully enclosing the oocyte within a continuous layer of follicle cells. At this moment, the egg becomes spatially complete. From now on, no cell will enter or leave.
FROM LIVING CELLS TO EGGSHELL
Once the oocyte is fully enclosed, the follicle cells shift roles yet again. Now they have become builders. The follicle cells begin secreting layers of proteinaceous material between themselves and the oocyte surface, forming the eggshell. This shell is not uniform: different regions of the follicle cell epithelium produce specialized structures, including:
The micropyle, a narrow opening for sperm entry
The operculum, a hatch through which the larva will emerge
The dorsal appendages, tube-like structures that enable gas exchange
Although the eggshell ultimately becomes an inert structure, its shape preserves the memory of follicle cell movements that preceded it. Morphogenesis is frozen into architecture.
Once the eggshell is complete, the egg chamber begins to dismantle itself. The nurse cells transfer the remainder of their contents to the oocyte and then degenerate. The follicle cells, having completed their task, also die off. What remains is a single egg—self-sufficient, protected, and ready for fertilization. The egg chamber, having fulfilled its purpose, ceases to exist.
CONCLUSION
The answer to the question “How does one egg chamber become one egg?” lies in precision and teamwork. Encapsulation isolates one germline cyst. Epithelial organization maintains strict boundaries. Morphogenetic movements reshape the chamber without cell division. Coordinated migration encloses the oocyte. Eggshell secretion replaces living tissue with structure. The egg chamber is not simply a container. It is a temporary organ, built to be dismantled, that exists solely to produce one egg.
Although fruit fly eggs may seem distant from human biology, the principles at work here are universal. Epithelial organization, cell polarity, coordinated migration, and tissue remodeling underlie the formation of organs throughout the animal kingdom. By understanding how one egg chamber becomes one egg, scientists gain insight into how complex structures are built and how they sometimes fail. In this way, the humble fruit fly egg chamber offers a powerful lesson: even the simplest biological outcomes can arise from astonishingly complex and beautiful processes.


Discussion